
Sexual Selection in Bowerbirds
Borgia Lab Web Site
|
|
|
|
Click here to see new videos |
Overview
We are interested in the evolution of mate choice and male
display in nonresource-based mating systems such as occurs in lekking
species. Most bowerbirds have a lek-like mating system and males
have no role in parental care but they build intricate and often finely crafted
stick structures associated with their display sites called bowers. We
study bowerbirds as a model system for understanding the evolution of complex
male sexual display and related processes like mate searching. Currently
we are engaged in a long term study of Satin Bowerbirds in northern NSW,
Australia but past studies have involved all other accessible species in this
family.
 |
Dr. Gerald Borgia | Graduate Students: | ||||||||||||||
Professor |
Joshua Kiner: jkiner001@gmail.com | |||||||||||||||
|
||||||||||||||||
Links:
Bowerbirds as a model system for understanding the evolution of complex male display
Current Research
Publications by the Borgia Lab
Information about volunteering as a field assistant to work in Australia
Information about volunteering to work as an undergraduate volunteer assistant in our lab
Information about applying as a graduate student to work in our lab
Videos
Information on sexual selection in individual bowerbird species (under construction)
Bowerbirds as a model system for understanding the evolution of complex male display.
The evolution of complex male display traits remains one of the most controversial issues in evolutionary biology. Bowerbirds provide an outstanding model system for testing hypotheses about the evolution and functional significance of complex display. This diverse group is best known for its polygynous species that build stick bowers on the ground associated with a decorated display; females provide parental care at nests built in trees. Multifaceted male displays involve the decorated display court and the bower that act as a stage for energetic vocal and dancing male display that is observed by the female from inside the bower. Courtship and matings occur in the bower. Because mating and courtship occur at a known location they can be monitored with video cameras and this has allowed the collection of highly detailed information on courtship and male mating success.
Bower quality, numbers of preferred decorations, and vocal/dancing elements all contribute to male mating success. Male reproduction is skewed; one male may mate with 25 different females at his bower in one season. Most females mate with a single male after visiting bowers of multiple males. Females having previously mated with high quality males show reduced mate searching and typically return to mate with these males in successive years.
Bowers likely originated as devices to protect females from forced copulation by courting males. For females to benefit from mate choice they must be able to visit and observe courtship by different males and reject those who are unsuitable. Female bowerbirds are susceptible to forced copulation when on the ground. The different bower types require the male to move around a barrier to copulate with the female and this allows disinterested females the opportunity to safely escape the display court. Archbolds bowerbird, a species that has lost bower- building, shows unique compensatory courtship displays in which males stay low and press their bodies close to the display court. Courting from this low position prevents the male from capturing the female from above as required for a forced copulation. Although, bowers reduce male opportunity for forced copulations, males with high quality displays likely gain from increased visitation of unfertilized females attracted to display courts that offer protection from forced copulation.
E. T. Gilliard suggested that in bowerbirds bright male plumage traits were transferred from the bird to the bower. This, however, is not supported in a phylogenetic analysis. An mt-DNA phylogeny shows that the monogamous catbirds are basal followed by separation of two clades that build distinctive bower types. Within each clade, bower form and decoration color use is highly variable. Reversals and convergences obscure the effects of shared ancestry and indicate no consistent trend from ancestral to derived species in bower and decoration complexity when contrasted with male plumage development. Results from several studies indicate that variation in bower form and decoration has arisen for different functional reasons.
Large changes in bowers allow improved conditions for the presentation of male display. In spotted bowerbirds, females prefer males giving high intensity displays but these displays are threatening. Changes in bower structure associated with high intensity display allow females to stand sideways in the bower and view male courtship displays protected by the see-though bower wall. Protection offered by the modified bower allows males to give higher intensity displays and ultimately more attractive displays. This threat reduction may be important in understanding display adaptations in other species. For example, male blue birds-of-paradise hang upside down in trees while displaying. This may have evolved as a device for reducing threat to females from high intensity displays.
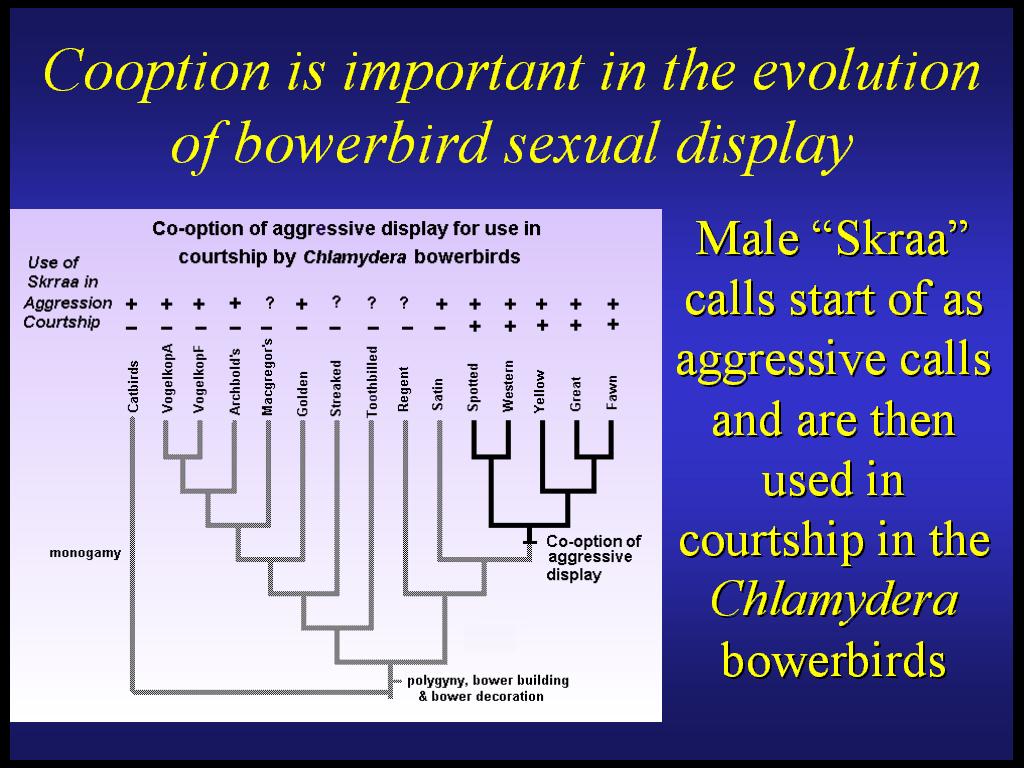
Some male display traits have been co-opted from other functions. Phylogenetic analysis of Skraa calls used across bowerbirds as aggressive calls suggests that they have been borrowed for use as courtship calls in the Chlamydera bowerbirds. Cooption of displays that signal male quality in aggressive contexts for use in courtship provides a simplified mechanism by which females can select for good genes. This does not require the difficult process of co-evolving male traits and female preferences required in most recent models. Bowers quality is also used in mate choice in one clade, but this is not a likely cause of bower evolution because incipient bowers would not reliably indicate quality differences among males. Similarly, decorations may have initially functioned as indicators of bower location and then been used in mate assessment. Thus, cooption and additions of secondary functions can have an important role in display trait evolution and can explain complex trait evolution without requiring coevolution of male traits and female preferences.
Males of different species choose colors of decorations in response to different light regimes. In Amblyornis bowerbirds, males in species using foggy ridge tops use predominantly black decorations while closely related species displaying on more sunlit slopes use a variety of decorations with a wide array of bright colors. The added cost of collecting bright decorations is justified on brightly lit slopes but may not be justified on poorly lit ridge top display sites. The effectiveness of functional explanations in describing shifts in bower structure and decoration provides an important alternative to runaway sexual selection as a cause of interspecific variation in display trait evolution.
Long term monitoring of satin bowerbird bowers shows that high male skew in mating success is maintained across multiple years causing variation in male lifetime reproductive success to be high. It has been suggested that costly male displays are necessary for honest advertisement of male quality, but the tendency for males to hold top reproductive positions for as long as six years suggests that display may not be extremely costly for them. Male bowerbirds do not obtain adult plumage until they are seven years and before then they frequently engage in male-male courtships where displays may be learned. Once acquired, male ability to display can be utilized in successive years at little additional cost. Recent studies suggest that attractiveness of male display comes from the intensity of display and from male ability to modulate display intensity in relation to female signals of comfort. Thus, in bowerbirds, experience and learning ability may insure quality signals rather than just difference in the costs of male traits that males can support and honest advertising need not be highly cost dependent.
Speculative discussions of bowerbirds and other species with elaborate display characterized males display as arbitrary, and tended to emphasize single models like runaway sexual selection. Information from detailed studies of bowerbirds suggest a different view. Male display evolved by a variety of processes. Rapid large changes in display can occur resulting in highly integrated displays meeting female needs, and are tuned to habitats occupied by individual species. While this adaptive view of mate choice has long been accepted in discussions of mate choice in species with resource-based monogamous mating systems, evidence suggests that it is now appropriate to extend it to species with nonresource-based mating systems that have elaborate male displays.


Current Research
Posters presented at recent scientific meetings. Please select title to see poster.
Co-option of pre-existing traits as a critical model for the evolution
of elaborate male sexual display
The Frequency of Use of a Cue Reflects Female Indecision in Mate Choice
Male intelligence influences male mating success in the satin bowerbird (Ptilonorhynchus violaceus)
Male aggression drives spatial association of kin in satin bowerbirds
Does cultural transmission of displays promote hybridization in bowerbirds?
Mismatch between high UV plumage reflectance and female preference for blue in satin bowerbirds
Experiments with robotic females show that successful males modulate courtship both spatially and temporally in response to signals of female discomfort in the satin bowerbird

The Wallaby Creek Field Site
Each year we recruit volunteer field assistants to help us with our studies
of satin bowerbirds in Australia.
The assistants' primary responsibility is
to help run cameras that are mounted at the bowers of each bird on
the study
site. Each assistant is responsible for maintaining 5-7 bowers and making
observations at these
bowers. The work includes carrying recharged car
batteries to bowers to power cameras. This is tough
and demanding work
that requires highly motivated and good natured assistants. There are
poisonous
snakes, land leeches and ticks on the study site and these are
potentially dangerous. We live in rustic
conditions and keep meal
preparation simple as most of our day is devoted to working on the birds.
The
great majority of assistants find the work demanding but rewarding and
there are many benefits to being at
Wallaby Creek, not the least being the
wide array of interesting wildlife. We begin recruiting assistants
in for work in the Fall (September through December). Assistants
are expected to pay their own way
to the field site. We house them and
provide food. Working as an assistant is an outstanding
opportunity to
learn about field work and is strongly encouraged for anyone interested in
applying as a
graduate student to work on bowerbirds in the Borgia
lab. This work is not for everyone, but if you are
interested
please contact Gerald Borgia.
Information about volunteering to work as an assistant in our lab
We return each year from Australia with more than 2000 Hi-8 video tapes that
were the result of
monitoring behavior at bowers. We recruit volunteer
assistants on the University of Maryland campus to
help us score
tapes. Students find this interesting because they can view the very
diverse behaviors of satin
bowerbirds and occasionally see a goana or python
crawl through the bower. Currently we do not
have funds to pay
volunteers, but qualified students can obtain credit for this work. It is
critical that
volunteers are highly accurate in their work. Work study
students are encouraged to apply.
If you are interested, please contact
Gerald Borgia.
Information
about applying to work as a graduate student in the Borgia Lab.
I encourage applications from students interested in sexual selection, the
evolution of display and related topics. All of the students in the lab are
working on bowerbird-related projects. This allows me to provide support
for students from my NSF awards. Opportunities exist for field work
and working in the laboratory.
Graduate students working in the
field are studying satin bowerbird evolution and behavior at our Wallaby Creek
Site. Current projects are on mate searching, juvenile development of male display traits, two-way signaling between males and
females in courtship, and studies of the relation between aerobic capacity
and male mating success. Central to all projects is a camera system that
monitors up 40 bowers continuously through the mating season. This
system combined with the large proportion of marked individuals in our
population produces an amazingly detailed data set that is useful for many
kinds of analysis. Using information available from video tapes students can test a wide array of hypotheses. Students working in the lab
work collaboratively with Mike Braun at the Laboratory of Molecular
Systematics (current project: Use of microsatellite markers to explore
relatedness), Rob Fleisher at the National Zoo (National
Zoo; current project: MHC variation, mate
choice and disease). The work at the field site is quite demanding and
difficult. Students lead a field crew of 6 volunteers over a four month
period. They must be scientifically astute, be able to deal with
rigorous conditions and effectively lead a diverse field crew. Ability to
carry out these functions and get along well with others in the lab is
important in assessing new students. I encourage students with high
GREs and grades to apply because they are eligible for fellowships that
provide support while they are in the field. Interested students should
contact me directly by email and provide grades, GRE scores, information about
relevant experience and references. Gaining familiarity with our
work by reading papers is strongly encouraged (see key downloadable papers below). Students may apply either to the BEES program or the Biology Department Graduate
Program at the University of Maryland. Contact Michelle Brooks at mmbrooks@umd.edu for application materials. |
Borgia, G. 1979. Sexual selection and the evolution of mating systems. In: Sexual
Selection and Reproductive Competition (Blum, M. & A., eds.) pp.
19-80. Academic Press. [pdf]
Alexander, R. D. and Borgia, G. 1979. On the origin and maintenance of the male-female
phenomenon. In: Sexual Selection and Reproductive Competition, Blum, M. & A.,
eds.) pp. 417-439. Academic Press.
[pdf]
Borgia, G. 1979. Levels of selection and human ethology. (Commentary) Behavioral and
Brain Sciences, 2: 30.
[pdf]
Borgia, G. 1980. Evolution of haplodiploidy: Models for inbred and outbred systems.
Theoretical Population Biology 17: 103-128.
[pdf]
Borgia, G. 1980. Sexual competition in Scatophaga stercoraria:
size- and density-related
changes in male ability to capture females. Behaviour 23: 185-206.
[pdf]
Borgia, G. 1980. Human aggression as a biological adaptation. In: The Evolution of Social
Behavior (Lockhard, J., ed.) pp. 165-191. Elsevier, North Holland.
Borgia, G. 1980. Review of Contrasts in Behavior. Q. Rev. Biol. 55: 95-96 [pdf]
Borgia, G. 1981. Mate selection in Scatophaga stercoraria: female choice in a
male-controlled system. Animal Behaviour 29: 71-80.
[pdf]
Borgia, G. and Blick, J. 1981. Sexual competition and the evolution of simultaneous
hermaphroditism. Journal of Theoretical Biology 84: 175-182.
[pdf]
Borgia, G. 1982. Experimental changes in resource structure and male density:
size-related differences in mating success among male Scatophaga stercoraria.
Evolution 36: 307-315.
[pdf]
Borgia, G. 1982. Female-biased sex ratios. Nature 298: 494-496.
Borgia, G. 1982. Experimental study of male choice in the satin bowerbird. American
Philosophical Society Yearbook pp. 24-25.
Borgia, G. 1982. Beyond sociobiology revisited. American Scientist. 70: 647-648.
Borgia, G. 1983. Reproductive behavior. Science 222: 920-922. [pdf]
Borgia, G. 1984. Insect sex. Bioscience 34: 524-525.
Borgia, G. 1985. Bowers as markers of male quality. Test of a hypothesis. Animal
Behaviour 33: 266-271
[pdf]
Borgia, G. 1985. Confidence of paternity and the avunculate. (commentary).
Behavioral and Brain Sciences 8: 68-89.
Borgia, G., Pruett-Jones, S. and Pruett-Jones, M. 1985. Bowers as markers of male
quality. Zietschrift fur Tierpsychologie 67: 225-236
[pdf]
Borgia, G. 1985. Bower destruction and sexual competition in the satin bowerbird
(Ptilonorhynchus violaceus). Behaviorial Ecology and Sociobiology18: 91-100.
[pdf]
Loffredo, C. and Borgia, G. 1986. Male courtship vocalizations as cues for mate
choice in the satin bowerbird (Ptilonorhynchus violaceus) Auk 103: 189-195.
[pdf]
Borgia, G. 1986. Satin bowerbird parasites: a test of the bright male hypothesis. Behaviorial
Ecology and Sociobiology 19: 355-358.
[pdf]
Borgia, G. and Gore, M. 1986. Sexual competition by feather stealing in the satin
bowerbird (Ptilonorhynchus violaceus). Animal Behaviour 34: 727-738.
[pdf]
Loffredo, C. and Borgia, G. 1986. Sexual selection, mating systems, and the
evolution of avian acoustical displays. American Naturalist 128: 773-794.
[pdf]
Borgia, G. 1986. Sexual selection in bowerbirds. Scientific American 254: 92 -101. [pdf]
Borgia, G., Kaatz, I. and Condit, R. 1987. Flower choice and the decoration of the bower of
the satin bowerbird (Ptilonorhyunchus violaceus): A test of hypotheses for the
evolution of display. Animal Behaviour 35: 129-1139.
[pdf]
Borgia, G. 1987. A critical review of sexual selection models. Dahlem Conference Symposium
Volume.Sexual Selection: Testing the Alternatives (Bradbury, J. and Andersson, M., eds.) pp.
55-66.Wiley: New York.
[pdf]
Heisler, L., Andersson, M., Arnold, S., Boak, C., Borgia, G., Hausfader, G., Kirkpatrick, M.,
Lande, R., Smith, J. Maynard, O'Donald, P., Thornhill, R. and Weissing, F. 1987.
Group report on models of sexual selection. Dahlem Conference Symposium
Volume. Sexual Selection: Testing the Alternative (Bradbury, J. and Andersson, M.,
eds.) pp. 98-118. Wiley: New York.
[pdf]
Borgia, G. and Collis, K. 1989. Female choice for parasite-free male satin bowerbirds and the
evolution of bright male plumage. Behavioral Ecology and Sociobiology 25: 445-454.
[pdf]
Borgia, G. 1989. Typology and human mating preferences. (commentary) Behavioral and
Brain Sciences. 12: 16-17.
[pdf]
Borgia, G. and Collis, K. 1990. Parasites and bright male plumage in the satin bowerbird
(Ptilonorhynchus violaceus). American Zoologist 30: 279-285.
[pdf]
Borgia, G. 1991. Sexual selection in Bowerbirds. In: Behavior and Evolution of Birds. D.
Mock, ed. New York: W.H. Freeman (Reprinting of 1986 Scientific American article).
[pdf]
Borgia, G. and Wingfied, J. 1991. Hormonal correlates of bower decoration and sexual display
in the satin bowerbird (Ptilonorhynchus violaceus). Condor. 93: 935-942
[pdf]
Borgia, G. and Mueller, U. 1992. Bower destruction, decoration stealing, and female choice in
the spotted bowerbird (Chlamydera maculata). Emu 92: 11-18.
[pdf]
Collis, K. and Borgia, G. 1992. The age-related effects of testosterone, plumage, and
experience on aggression and social dominance in juvenile male satin bowerbirds
(Ptilonorhynchus violaceus). The Auk. 109: 422-434.
[pdf]
Borgia, G. and Wilkinson, G. 1992. Swallowing ornamental asymmetry. Nature 359: 487-488. [pdf]
Borgia, G. 1993. The cost of display in the non-resource based mating system of the satin
bowerbird. American Naturalist. 141: 729-743.
[pdf]
Collis, K. and Borgia, G. 1993. Delayed plumage maturation, facilitated learning, and the cost
of display in the satin bowerbird: An experimental study. Ethology. 94: 59-71.
[pdf]
Kusmierski, R., Borgia, G., Crozier, R. and Chan, B. 1993. Molecular information on
bowerbird phylogeny and the evolution of exaggerated male characteristics. Journal of
Evolutionary Biology. 6: 737-752.
[pdf]
Borgia, G. 1994. The Scandals of San Marco. Quart. Rev. Biol. 69: 373-377. [pdf]
Borgia, G. 1995. Why do bowerbirds build bowers? Amer. Scientist. 83: 542-547. [pdf]
Borgia, G. 1995. Treat reduction as a cause for large differences in bower architecture, in two
closely related bowerbird species Chlamydera nuchalis and C. maculata. Emu
95: 1-12.
[pdf]
Borgia, G. 1995. Complex male display and female choice in the spotted bowerbird:
Specialized functions for different bower decorations. Animal Behaviour 49: 1291-1301.
[pdf]
Frith, C., G. Borgia, D. Frith. 1996. Courts and courtship in Archbold's Bowerbird,
Archboldia papuensis (Ptilonorhynchidae) in Papua New Guinea. Ibis
138: 204-211.
[pdf]
Borgia, G. 1996. Comparative behavioral and biochemical studies in bowerbirds and
the evolution of bower building. In Biodiversity II. Eds. M. Reaka, D.
Wilson, and E.O. Wilson, Smithsonian Institution, Chapter 18 pp. 246-263.
Washington, D.C.
[pdf]
Borgia, G. 1996. Satin bowerbird displays are not extremely costly. Animal Behavior 52: 221-224. [pdf]
Borgia, G. 1996. Sexual warfare. Science 272: 1723. [pdf]
Kusmierski, R., G. Borgia, A. Uy, and R. Crozier. 1997. Molecular information on bowerbird phylogeny
and the evolution of exaggerated male characters. Proc. R. Soc. Lond. 264, 307-313.
[pdf]
Borgia, G. 1998. Why do bowerbirds build bowers? Ch 17. In: Exploring Animal Behavior.
Edited by Paul Sherman and J. Alcock, Sinauer Associates, Inc.
[pdf]
Borgia, G. and D. Presgraves, 1998. Coevolution of elaborated male display traits in the
spotted bowerbird: An experimental test of the threat reduction hypothesis. Animal
Behavior, 56: 1121-1128.
[pdf]
Uy, A., Patricelli, G. and Borgia, G. 2000. Dynamic mate searching tactic allows female satin
bowerbirds to reduce searching. Proc. R. Soc. Lond. B. 267: 251- 256.
[pdf]
Uy, J.A.C. and Borgia, G. 2000. Sexual selection drives rapid divergence in bowerbird display
traits. Evolution. 54(1): 273-278.
[pdf]
Borgia, G and Coleman, S. 2000. Co-option of male courtship signals from aggressive
display in bowerbirds. Proc. R. Soc. Lond. B. 267: 869-874.
[pdf]
Uy, J. A. C., G. Patricelli and G. Borgia. 2001. Loss of preferred mates forces female satin bowerbirds
(Ptilonorhynchus violaceus) to increase mate searching. Proc. R. Soc. Lond. B. 268: 633-638.
[pdf]
Uy J.A.C., Patricelli G.L, Borgia G. 2001. Complex mate searching in the satin bowerbird
Ptilonorhynchus violaceus American Naturalist 158 (5): 530-542.
[pdf]
Borgia, G. 2002. Sexual selection: Bowerbirds. Encyclopedia of Evolution. M. Pagel Ed.
Oxford University Press, pp. 1051-1053.
Patricelli G.L., Uy J.A.C., Walsh G. & Borgia G. 2002. Sexual selection: males adjust displays
in response female signals. Nature 415: 279-280.
[pdf]
Patricelli G.L., Uy, J.A.C., Borgia G. 2003. Multiple male traits interact: attractive bower decorations
facilitate attractive behavioural displays in satin bowerbirds. Proc. R. Soc. Lond., B 270, 2389-2395.
[pdf]
Coleman, S.W., Patricelli, G.L., Borgia, G. 2004. Variable female preferences drive complex male
displays.
Nature 428: 742-745.
[pdf]
Patricelli, G.L., Uy, J.A.C., & Borgia, G. 2004. Female signals enhance the efficiency of mate assessment
in satin bower-birds (Ptilonorhynchus violaceus). Behav. Ecol. 15 (2): 297-304.
[pdf]
Borgia, G., M. Egeth, J. A. C. Uy, G. L. Patricelli. 2004. Juvenile infection and male display: testing
the bright male hypothesis across individual life histories. Behav. Ecol. 15: 722-728.
[pdf]
Borgia, G. 2004. Reproductive behavior: Bowerbirds and sexual displays. pp 881-883.
Marc Bekoff ed. Encyclopedia of Animal Behavior. Greenwood Press, London.
Shaw, K., Mendleson, T, and Borgia, G. 2005. Evolution by sexual selection.
In Evolutionary Science and Society. pp. 99-108, Cracraft, J. and Bybee, R. eds. AIBS: Washington D.C.
Borgia, G. 2005. The Bowerbirds: Ptilonorhynchidae. The Auk 122: 718–721, 2005 [pdf]
Patricelli GL, Coleman SW, Borgia G. 2006. Male satin bowerbirds, Ptilonorhynchus violaceus,
adjust their display intensity in response to female startling: an experiment with robotic females Anim. Behaviour 71: 49-59
[pdf]
Borgia, G. and J. Keagy. 2006. An inverse relationship between decoration and food
colour preferences in satin bowerbirds does not support the sensory drive hypothesis, Animal Behaviour, 72 : 1125-1133.
[pdf]
Borgia, G. 2006. Preexisting Male Traits are Important in the Evolution of Elaborated
Male Sexual Display. Advances in the study of Behaviour 36: 249-302.
[pdf]
Reynolds, S.M., Dryer, K, Bollback, J. Uy, JAC,Patricelli, GL, Robson, T, Borgia, G., & Braun,
M.J. 2007. Behavioral paternity predicts genetic paternity in satin bowerbirds (Ptilonorhynchus violaceus),
a species with a non-resource-based mating system. The Auk 124: 857867.
[pdf]
Borgia, G. Coyle, B. & Zwiers, P. 2007. The Evolution of Colorful Display. Evolution 61(3): 708-12 [pdf]
Coleman SW, Patricelli GL, Coyle B, Siani J, and Borgia G. 2007. Female preferences drive the
evolution of mimetic accuracy in male sexual displays. Biol. Lett. 3, 463466.
[pdf]
Zwiers PB, Borgia G, Fleischer RC. Plumage based classification of the Bowerbird genus Sericulus evaluated using a multi-gene, multi-genome analysis. Molecular Phylogenetics and Evolution. Mar;46(3):923-31. Epub 2007 Dec 5 [pdf]
Borgia, G. 2008. Blocking of UV reflectance does not influence use of off-body
display elements by satin bowerbirds. Behavioral ecology 19 (4) 740-746. (doi:10.1093/beheco/arn010)
[pdf]
Reynolds, SM, Christman MC, Uy JAC, Patricelli GM, Braun MJ, and Borgia G. 2009. Lekking satin bowerbird males aggregate with relatives to mitigate aggression. Behavioral Ecology 20, 410-15. (doi:10.1093/beheco/arn146) [pdf]
Keagy, J., Savard, J-F, and Borgia, G. 2009. Male satin bowerbird problem-solving ability predicts mating success. Animal Behaviour. 78: 809-817. [pdf]
Keagy, J, Savard, JF, Borgia, G. 2011. Complex relationship between multiple measures of cognitive ability and male mating success in satin bowerbirds, Ptilonorhynchus violaceus. Animal Behaviour 81: 1063-1070. [pdf]
Savard, JF, Keagy, J, Borgia, G. 2011. Blue, not UV, plumage color is important in satin bowerbird Ptilonorhynchus violaceus display. Journal of Avian Biology 42: 80-84.
Chappell, MA, Savard, JF, Siani, J, Coleman, SW, Keagy, J, Borgia, G. 2011. Aerobic capacity in wild satin bowerbirds: repeatability and effects of age, sex and condition. The Journal of Experimental Biology 214: 3186-3196. [pdf]
Borgia, G., Coyle, B. and Keagy, J. 2012. Comment on "Illusions Promote Mating Success in Great Bowerbirds". Science. 337: 292-292. [pdf]
Keagy, J., Savard, J-F, and Borgia. G. 2012. Cognitive ability and the evolution of multiple behavioral display traits. Behavioral Ecology. 23: 448-456. [pdf]
Coyle B J, Hart N S, Carleton K L, & Borgia G 2012. Limited variation in visual sensitivity among bowerbird species suggests that there is no link between spectral tuning and variation in display colouration. J Exp Biol 215:1090-1105. [pdf]
Hicks*RE, Larned A, Borgia G. 2013. Bower paint removal leads to reduced female visits, suggesting bower paint functions as a chemical signal. Animal Behaviour 85:1209-1215. [pdf]
Borgia, G. and Keagy, J. 2015. Sexual selection and cognitive ability: what bowerbirds can teach us. In Animal Signaling and Function: An Integrative Approach. D. Irschick, M. Briffa, J. Podos, Eds. John Wiley and Sons. [pdf]
Keagy, J., Hosler, L.C*, and Borgia, G. 2016. Female active sampling of male paint on bowers predicts female uncertainty in mate choice. Animal Behaviour. 116: 131-137. [pdf] featured in that issue's In Focus section [pdf]
Borgia, G. and Ball G. 2017. The evolution of beauty: how Darwin's forgotten theory of mate choice shapes the animal world - and us, Richard O. Prum. New York: Doubleday (2017). 448 pp. [pdf]
