
Non-electrolytes:
Rather large non-electrolytes are able to permeate through VDAC channels (refs. 1-3):
-polyethylene glycols up to a molecular weight
of 3400
-inulin
-gamma cyclodextrin
-dextrans up to a molecular weight of about 5000
Ionized atoms and molecules:
Channel permeability to charged species is strongly dependent on
the sign of the charge. In its highest conducting state, the open
state, there is a strong preference for anions (see table, refs.
5-7). This preference becomes inverted upon channel closure, conversion
to a low-conducting state. This can be understood in terms of the
gating model by which a positively-charged domain moves from within
the channel (forming a portion of the wall of the channel) to the
membrane surface.
single channel flux (ions/msec)
| State | K+ | Cl- | succinate2- | citrate3- | HPO42- | ATP4- | radius (nm) |
| open | 13 | 67 | 6.8 | 3.4 | 6.3 | 1.1 | 1.4 |
| closed | 26 | 4.5 | 0.3 | 0.2 | 0.3 | 0.0 | 0.9 |
Small Ions:
The selectivity for small ions, as measured by the reversal potential in the presence of a salt gradient, can be understood rather well in terms of simple electrostatics within the pore. Using a modification of Teorell’s theory for ion flow through an ion exchange membrane, a "Large Channel Theory" (LCT) was developed (ref. 4). It can account rather well for the observed reversal potential under a variety of conditions using only one adjustable parameter (the charge on the wall of the pore). Even this parameter conforms closely to experimental measurements.
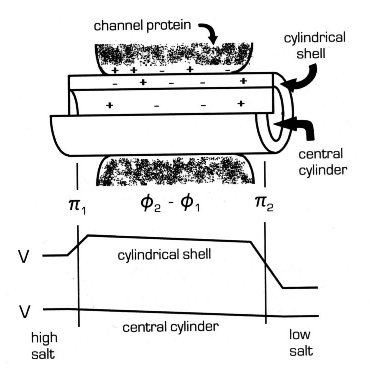
Schematic representation
of the partitioning of the queous pore into two compartments and
the electrical potential profile in each. The net charge on
the inner wall of the channel results in a cylindrical layer of
charged solution next to the protein (the cylindrical shell).
The thickness of this shell is proposed to be a mean of the critical
distances calculated for the ionic strength at each end of the channel.
The central cylinder is therefore relatively neutral. The
cylinders of solution extend beyond the end of the protein because
of access resistance regions. The electrical potential
changes along the cylindrical shell are the two Donnan potentials
at the ends of the pore (p1 and p2)
and
the diffusion potential (f1 - f2 ) in the pore.
Donnan potentials do not apply to the central cylinder.
LCT proposes that the aqueous compartment within the channel can be divided into 2 regions: a cylindrical shell of solution next to the charged inner wall of the channel and a cylindrical central region that is essentially filled with bulk salt solution. The cylindrical shell is considered to be a fixed charge region whose ionic composition is determined by the proximity of the fixed charges on the wall of the pore. The thickness of the region is determined by the distance required for the potential to reach a value that is 1/e of that at the surface of the protein. It is equivalent to the Debye length but in cylindrical coordinates.
Despite its simplicity, it can account for:
1. the biphasic relationship between the reversal potential and the transmembrane salt activity ratio
2. the dependence of the reversal potential on ionic mobility
3. the dependence the reversal potential on charge in the pore as altered by site-directed mutations
and to a lesser extent the dependence of the reversal potential on salt activity (keeping the activity ratio constant).
Simple Goldman/Hodgkin/Katz treatment does not even come close. Other approaches require the adjustment of free parameters to begin to reflect the observations. Despite its simplicity, LCT is the theory that shows the best agreement with the experimental observations.
Organic Ions:
Unlike small ions, organic ions may interact more specifically with the specific location of the charges on the wall of the channel. Evidence exists for an ATP binding site in VDAC (refs. 7,8). The gating of VDAC converts the structure of the channel from one that allows the passage of ATP to one that does not (Figure).
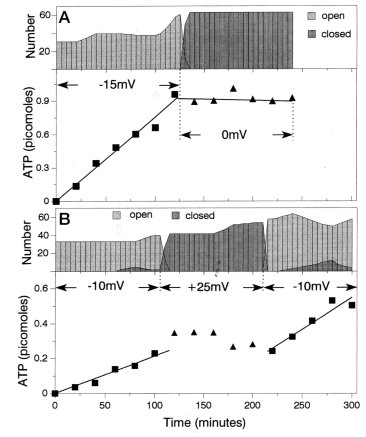
The ATP flux through
the open and closed states of VDAC channels. Two experiments
are illustrated: one in A (cis [ATP] = 153 mM), the other in B (cis
[ATP] = 60 mM). The lower portions of A and B show the amounts
of ATP in the trans compartment as a function of time. The
indicated voltage was applied to control the state of the channel.
The squares show samples collected while the channels were open,
and the triangles when the channels were closed. The upper
portions show the number of open and closed channels at any time
during the experiment. These were determined from continuous
measurements of conductance and selectivity throughout the experiment
using the single-channel characteristics obtained from parallel
single-channel experiments.
Selectivity and Channel Gating:
While channel closure does result in a reduction in effective pore radius (from non-electrolyte measurements), the major impact in a alteration in selectivity stemming from a radical change in the net fixed charge lining the walls of the pore. The combination of a reduction in pore size and inversion in selectivity can cause an actual increase in K+ flow (see table) but a drastic decrease in the flow of anions. It must be emphasized that cation and anion flows are linked and so the measured value of one depends on the presence of the other. Thus the numbers in the table must be used with caution but the main message, regarding the profound effect of the selectivity change, is clear.
References:
1. Colombini, M. 1980. Structure and mode of action of a voltage-dependent anion-selective channel (VDAC) located in the outer mitochondrial membrane. Annals of the New York Academy of Sciences, 341: 552-563.
2. Colombini, M. 1980. The pore size and properties of channels from mitochondria isolated from Neurospora crassa. Journal of Membrane Biology, 53: 79-84.
3. Zalman, L.S., Nikaido, H., and Kagawa, Y. 1980. Mitochondrial outer membrane contains a protein producing non-specific diffusion channels. J. Biol. Chem. 255:1771-1774.
4. Zambrowicz, E.B. and Colombini, M. 1993. Zero-current potentials in a large membrane channel: a simple theory accounts for complex behavior. Biophysical Journal, 65: 1093-1100.
5. Rostovtseva, T. and Colombini, M. 1996. ATP flux is controlled by a voltage-gated channel from the mitochondrial outer membrane. Journal of Biological Chemistry, 271: 28006-28008.
6. Hodge, T. and Colombini, M. 1997. Regulation of metabolite flux through voltage-gating of VDAC channels. Journal of Membrane Biology, 157: 271-279.
7. Rostovtseva, T. and Colombini, M. 1997. VDAC channels mediate and gate the flow of ATP: implication on regulation of mitochondrial function. Biophysical Journal, 72: 1954-1962.
8. Rostovtseva, T. and Bezrukov, M. 1998. ATP transport to a single
mitochondrial channel, VDAC, studied by current fluctuation analysis.
Biophysical Journal, 74: 2365-2373.
|
Please send comments
and contributions to Dr.
Marco Colombini
|
|
Last modified: March 27, 2004 15:32 Page created and maintained by Swapnil Sharma |